Can Handedness Be Determined From Skeletal Remains? A Chronological Review of the Literature
![]()
Biomechanical Evaluation on the Bilateral Asymmetry of Complete Humeral Diaphysis in Chinese Archaeological Populations
1
Cardinal Laboratory of Vertebrate Evolution and Human being Origins, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese University of Sciences, Beijing 100044, Communist china
2
CAS Middle for Excellence in Life and Paleoenvironment, Beijing 100044, China
3
College of Globe and Planetary Sciences, University of Chinese University of Sciences, Beijing 100049, Cathay
4
Plant of Archeology and Cultural Relics of Hubei Province, Wuhan 430077, China
5
Schoolhouse of Ethnology and Sociology, Minzu Academy of China, Beijing 100081, China
6
Heart for the Study of Chinese Archeology, Peking University, Beijing 100871, Red china
seven
Ministry of Education Cardinal Laboratory of Contemporary Anthropology, Department of Anthropology and Homo Genetics, School of Life Sciences, Fudan University, Shanghai 200433, Cathay
8
Middle for the Exploration of the Deep Human being Journey, Faculty of Science, University of the Witwatersrand, 2050 Johannesburg, South Africa
*
Author to whom correspondence should be addressed.
Academic Editor: Antoine Balzeau
Received: 28 Baronial 2021 / Revised: 26 September 2021 / Accepted: 28 September 2021 / Published: 2 October 2021
Abstract
Diaphyseal cross-sectional geometry (CSG) is an effective indicator of humeral bilateral asymmetry. However, previous studies primarily focused on CSG backdrop from limited locations to represent the overall bilateral biomechanical operation of humeral diaphysis. In this study, the complete humeral diaphyses of 40 pairs of humeri from three Chinese archaeological populations were scanned using loftier-resolution micro-CT, and their biomechanical asymmetries were quantified by morphometric mapping. Patterns of humeral disproportion were compared between sub-groups defined by sex and population, and the representativeness of torsional rigidity asymmetry at the 35% and 50% cantankerous-sections (J35 and Jfifty asymmetry) was testified. Inter-group differences were observed on the hateful morphometric maps, only were non statistically significant. Analogous distribution patterns of highly asymmetrical regions, which correspond to major muscle attachments, were observed across near all the sexes and populations. The diaphyseal regions with high variability of bilateral asymmetry tended to present a depression asymmetrical level. The J35 and J50 asymmetry were related to the overall humeral disproportion, but the correlation was moderate and they could not reverberate localized asymmetrical features beyond the diaphysis. This written report suggests that the overall asymmetry pattern of humeral diaphysis is more complicated than previously revealed by individual sections.
1. Introduction
Humeral bilateral asymmetry has been extensively studied in orthopedics, forensics, and paleo/archaeological anthropology [1,2,3]. Handedness can be inferred from the bilateral asymmetry of the upper limb [four,5,6]. Evidence from living athletes of unilaterally dominated sports (such equally lawn tennis and cricket) suggests a close relationship between humeral bilateral disproportion and behavioral laterality [seven,8,9]. A combined written report of endocranial and humeral asymmetry tin can shed light on how the human body responds to dependent asymmetrical stimuli across biologically independent anatomical regions [10]. These applications make humeral bilateral asymmetry an effective approach for reconstructing the behaviors of by human populations [11,12,13,14,fifteen,sixteen].
Long os diaphyses show great plasticity to remodel in response to mechanical loadings beyond a lifetime, especially prior to sexual maturity [17,18,19,20,21]. This remodeling makes diaphyseal cross-sectional geometry (CSG) a more effective indicator of bilateral upper-limb use and asymmetry compared to other linear measurements, such equally articular jiff or bone length [13,20,22,23,24].
Polar moment of area (J) and 2nd moment of area (SMA) are 2 normally adopted CSG parameters in humeral biomechanical analysis. J indicates the cantankerous-section's torsional and average bending rigidity, whereas SMA denotes the verbal angle rigidity along a sure axis of a cross-section [3,25]. Owing to the difficulties of obtaining sequential histological cross-sections, nearly earlier studies focused on the CSG properties of cross-sections placed at 35% or 50% of the humeral biomechanical length (see the definition made past Ruff [26]). J at the 35% cross-department (J35) can reasonably estimate the minimum rigidity of humeral diaphysis and avoids the interference of other anatomical features, every bit it is situated below the distal edge of deltoid tuberosity and above the supracondylar crest [13,fourteen,16,27,28,29]. J at the 50% cross-department (J50) provides reasonable estimates of midshaft rigidity [ix,14,30,31,32,33,34], and is known to exist a reliable indicator of hand preference [5]. When evaluating the directional biomechanical performance of a cross-section, most previous studies only calculated the maximum/minimum SMA or SMA forth the standard anatomical centrality (anteroposterior or mediolateral) to avoid the complexity of acquiring CSG values in multiple directions [35,36,37].
However, CSG properties of limited cross-sections and directions are bereft to gauge the overall biomechanical operation of long os diaphysis, especially in studies near humeral bilateral disproportion. According to experimental data from professional baseball players, tensile and shear strains vary among different diaphyseal sections during throwing activities [38], and the degree of bilateral disproportion evaluated by J was variable forth the humeral shaft [16,38]. The shape asymmetry of different cantankerous-sections also indicates that the asymmetry patterns vary in dissimilar anatomical directions [24,39].
Morphometric mapping is a 2D visualizing method that is ordinarily used for displaying the distribution patterns of morphometric and biomechanical properties across the entire diaphysis of a long bone [40,41]; for example, the distribution patterns of cortical bone thickness along the femoral diaphysis, visualized by morphometric maps, differentiate in Neanderthals and Homo erectus from mod humans [42,43]. Additionally, morphometric maps, quantifying the external radius across the entire femoral diaphysis, reveal the ontogenetic disparities between wild and captive chimpanzees [44]. The cortical structure of hallucal metatarsals, represented past morphometric maps of cortical bone thickness and bending rigidity, reflects locomotor adaptations of humans, chimpanzees, and gorillas [45]. Finally, morphometric mapping has been established to exist an effective approach for quantifying the humeral biomechanical disproportion across the complete diaphysis [sixteen].
Factors such every bit geographic location, chronological historic period, subsistence pattern, and sex are known to influence the design of humeral asymmetry in man populations. Varying degrees of humeral disproportion have been detected amid Upper Paleolithic populations from Europe, Africa, and Asia [11]. European samples prove a general subtract in humeral asymmetry from the early Upper Paleolithic populations through to the 20th century [13,22]. Foragers and farmers from the pre-Hispanic American Southwest present different humeral asymmetry patterns [36]. Due to the beingness of the sexual division of labor, modern homo populations with various geographic locations, chronological ages, and subsistence patterns tend to exhibit diverse sexual dimorphism patterns in humeral asymmetry [31,36,37,46].
In the present study, we aim to (1) generate a more than comprehensive understanding of humeral disproportion past evaluating the biomechanical performance across complete diaphysis compared to previous studies, which only used individual cross-sections; and (ii) cheque the reliability of using J35 and J50 to represent the overall humeral biomechanical performance in bilateral asymmetry analysis. To fulfil these targets, specimens were scanned using loftier-resolution micro-computed tomography (micro-CT), and morphometric mapping was practical to quantify the overall biomechanical disproportion of humeral diaphysis for its effectiveness in visualization and statistical analysis. To cover as broad a variety of specimens as possible, xl pairs of humeri from 3 Chinese archaeological populations, which differ in geographic location, chronological historic period, and subsistence strategy, were selected to correspond E Asian modern humans in the present written report.
ii. Materials and Methods
2.one. Materials
Forty pairs of modernistic human humeri are included in this written report. All paired humeri were collected from archaeological sites with populations that varied in geographic location, chronological historic period, and subsistence design. Agronomical and nomadic/gathering populations are included because these lifestyles were the dominant subsistence patterns in pre-industrial Eastern asia. The subsistence patterns of these populations were determined by associated burying assemblages and relevant historical records. The populations volition exist referred to by their geographic locations, which are as follows: ①Hubei population (HB): nine males and iv females nerveless from agricultural sites from Hubei Province, Central People's republic of china spanning Qin-Han-Tang dynasties (221 BC ~ 907 Advertising). For some sites of this population, analyses of charred institute remains indicate that Setaria italica and Panicum miliaceum were the primary nutrient crops [47]. ②Henan population (HN): vi males and v females collected from an agricultural population from Junzicun cemetery, Henan Province, North Red china, which dates to Qing dynasty (1636 Advertisement ~ 1912 Advertising). Historical records signal that an agronomical economy was the dominant lifestyle of this population [48]. ③Xinjiang population (XJ): 10 males and 6 females collected from nomadic populations attributed to Subeixi civilisation (yard BC ~ 200 BC) from the Turpan Basin, Xinjiang Province, Northwest China. Burial assemblages such as bows, arrows, and stone artifacts for males and spinning wheels and potteries for females indicate a subsistence pattern of nomadism and gathering [49,50,51]. All individuals were adults. Their historic period and sex activity were determined according to cranial and pelvic osteological indicators. All humeral specimens were intact, well preserved, and showed no symptoms of osteoporosis or other pathologies.
2.2. Data Collecting and Processing
All humeri were scanned past a 450 kV micro-CT scanner (designed past Institute of High Free energy Physics, Chinese Academy of Sciences) located in Key Laboratory of Vertebrate Development and Human Origins, Constitute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences. The scanning was performed under a voltage of 380 kV, electric current of 1.5 mA, 360° rotation with a step of 0.5°, and an isometric voxel size of 160 μm. Raw information were virtually reconstructed and segmented in VGStudio Max iii.0. Book renderings of all humeri were aligned to anatomical position using the standard protocol defined by Ruff [26]. To ensure that the humeri were consistently aligned and to avoid inter-observer error, all alignments were made by one author (Y.Z.). Paired humeri were always aligned synchronously. 3-dimensional meshes of each aligned humerus were generated and saved as PLY files in Avizo 8.one for the following analyses.
2.3. Cross-Exclusive Geometric Parameters Calculation
Customized in-firm scripts, mainly sourced from R package 'morphomap', were applied to calculate the CSG parameters [52]. For each humerus, the unmarried-layer periosteum and endosteum surface meshes were firstly discrete from the original humeral mesh. Second, 61 equidistant cross-sections were extracted from the surface meshes along the proximodistal diaphysis (betwixt 20 and fourscore% of the biomechanical length). 3rd, 360 equiangular landmarks were placed forth both the inner and outer contours on each cross-section. Finally, J values of the cantankerous-sections at 35% and 50% of biomechanical length (J35 and J50), and SMA values of 360 directions on 61 cantankerous-sections were calculated based on the landmark coordinates.
2.four. Bilateral Disproportion Quantification
Ordinarily used practices for assessing bilateral asymmetry are absolute asymmetry ([(max − min)/((max + min)/ii)] × 100%) and directional asymmetry ([(right − left)/((correct + left)/2)] × 100%). However, absolute asymmetry is non appropriate in this study, equally the magnitude relationship between the left and right side is not consistent among different landmarks at humeral diaphysis. Even so, our study nonetheless focuses on absolute information of overall bilateral disproportion, then directional asymmetry is also not suitable, because it does not eliminate the impact of handedness as well as behavioral laterality, which is non the issue this study attempts to investigate and may bring about bias to the determination. Therefore, bilateral disproportion was quantified using dominant asymmetry ([(dominant − non-dominant)/((dominant + non-ascendant)/2)] × 100%). The dominant side was decided according to the magnitude of J50, given that it is a valid indicator of handedness [5].
2.5. Morphometric Mapping
The SMA asymmetry values were obtained using the dominant asymmetry equation for all 21,960 (360 × 61) landmarks, and the results for each paired humeri were deposited in a matrix with 61 rows (sorted by the guild of cross-sections) and 360 columns (sorted by the order of directions). These matrices were then visualized every bit morphometric maps to brandish the distribution characteristics of bending rigidity asymmetry along the proximodistal humeral diaphysis (Figure one and Figure A1). The asymmetry values of J35 and Jl for all individuals were also calculated using the aforementioned equation.
2.6. Methods to Estimate the Variation of Humeral Biomechanical Asymmetry
To explore the variation in humeral asymmetry patterns in modern humans, twoscore individuals were divided into sub-groups defined by sexual activity and population. The iii populations, which varied in geographic location, chronological historic period, and subsistence blueprint, were supposed to vary in their habitual behaviors, and then population was set up every bit one variable. Sexual division of labor is an important effect when discussing historical populations, and the sexual dimorphism of humeral asymmetry can be affected by non-behavioral factors such as genetics or hormones [27]. Therefore, sexual activity was set equally some other variable. Mean morphometric maps exhibiting SMA asymmetry values for each sub-grouping were qualitatively compared. Additionally, a two-way multivariate analysis of variance (MANOVA) was conducted to quantitatively test whether sex and/or population were significant sources of variation. When plumbing equipment the regression model for MANOVA, SMA disproportion values at all landmarks were set every bit the dependent variables, while sex and population were set up as the independent variables with interaction. Customized in-house scripts, mainly sourced from R package 'geomorph' and 'RRPP', were utilized to bear MANOVA [53,54]. In addition, the coefficients of variation (CV) for SMA asymmetry values at all landmarks were calculated in sub-groups and visualized by morphometric maps to showroom intra-group variation characteristics. Only sub-groups divers past sex or by population were included in this assay to reduce the impact of outliers.
ii.7. Methods to Exam the Representativeness of J35 or J50 Asymmetry
The reliability of using J35 or Jfifty asymmetry to stand for the overall humeral asymmetry was tested using several statistical methods. First, a multivariate regression model was congenital on all specimens to statistically examination the degree of correlation betwixt overall SMA disproportion and J asymmetry. When fitting the model, the SMA asymmetry values at all landmarks were set up as the dependent variables, and the J35 or J50 disproportion value every bit the contained variable. Customized in-house scripts, mainly sourced from R packet 'geomorph' and 'RRPP', were utilized to carry out this fitting [53,54]. Second, to investigate the clan of every SMA disproportion value and J disproportion value across the entire humeral diaphysis, the correlation coefficients between each SMA asymmetry value and J35 or Jl asymmetry value (CC35 and CC50) were calculated within sub-groups. The same protocols for visualizing SMA asymmetry values were applied to CC results to generate morphometric maps. The CC morphometric maps of sub-groups were qualitatively compared to reveal inter-group variations.
3. Results
3.1. Pattern of Humeral Biomechanical Disproportion in Mod Humans
The mean morphometric maps exhibiting SMA asymmetry values for each sub-grouping and pooled samples are presented in Figure 2. Hubei females and males are more asymmetrical in the near-anterolateral posteromedial aspect forth the unabridged proximodistal diaphysis. The degree of asymmetry is transversely uniform around the mid-distal diaphysis for Hubei females, and around the midshaft for Hubei males. Hubei males accept higher anteroposterior asymmetry from the proximal to mid-proximal diaphysis. Henan females have a restricted area of relatively higher anteroposterior asymmetry around the mid-proximal diaphysis, while Henan males are more asymmetrical in the nearly-anterolateral posteromedial aspect spanning the mid-proximal to distal diaphysis. Both Xinjiang females and males accept reinforced anteroposterior asymmetrical areas effectually the proximal diaphysis, as well equally the region between the proximal to mid-proximal diaphysis, mediolaterally. The region with a relatively college asymmetry of Xinjiang males extends from the midshaft to the distal diaphysis in the virtually-anterolateral posteromedial aspect.
For the mean morphometric maps that are defined only past population, Hubei is more asymmetrical across the entire proximodistal diaphysis in the nigh-anterolateral posteromedial attribute, with a reinforcement of anteroposterior asymmetry forth the proximal to mid-proximal diaphysis. The region with high disproportion for Henan is located in the anterolateral posteromedial aspect between the mid-proximal to distal diaphysis. Xinjiang has higher anteroposterior asymmetry around the proximal diaphysis, connecting with some other area with high mediolateral disproportion effectually the mid-proximal diaphysis, which continuously extends to the midshaft in the anterolateral posteromedial aspect. Hubei and Xinjiang are more asymmetrical than Henan, according to their overall magnitude of SMA asymmetry values. For the mean morphometric maps that are defined only by sex, females are more anteroposteriorly asymmetrical between the proximal and mid-distal diaphysis, with a reinforcement of asymmetry near the mid-proximal section. The distribution patterns of males resemble that of Xinjiang, simply the regions with highest asymmetry at the proximal and mid-proximal diaphysis are not and so prominent, and the region with relatively higher asymmetry along the distal half of the diaphysis in the anterolateral posteromedial aspect is more developed. Males are more asymmetrical than females in general. The mean morphometric map for pooled samples shows uniform areas of asymmetry spanning from the proximal diaphysis, anteroposteriorly, to the mid-proximal diaphysis, mediolaterally, and standing distally in the anterolateral posteromedial aspect.
Co-ordinate to the results of MANOVA (Table 1), the differences sourced from sex (P = 0.11), population (P = 0.296), and the interaction of sex activity and population (P = 0.783) are not statistically significant. The R-squared values reveal that sex activity, population, and the interaction of sex and population accounted for 5.49%, 5.99%, and 2.74% of the total variation, respectively. Residuals accounted for 85.77% of the total variation.
The CV morphometric maps show about identical distribution patterns across all the sub-groups and pooled samples (Figure 3). Relatively high CV values are concentrated in the region between the heart and mid-distal diaphysis, and at the distal extreme in the anteromedial posterolateral aspect. Similarly loftier CV values announced at the proximal department, mediolaterally, merely to a smaller extent compared to the distal department. Henan has localized regions of higher CV values at the proximal extreme, mediolaterally, and at the mid-distal diaphysis in the anteromedial posterolateral aspect, but displays no other differences compared to Hebei and Xinjiang. Females present higher overall CV values than males.
3.2. Representativeness of J35 and Jl Bilateral Asymmetry
Table 2 and Figure 4 show the result of a multivariate regression fitting all the SMA asymmetry values on the J35 or J50 asymmetry value using pooled samples. The results of J35 and J50 asymmetry are highly significant (P < 0.001), indicating that the multivariate regression model is constructive. According to the R-squared values, J35 asymmetry accounts for 48.66% of the full variation, whereas Jfifty asymmetry accounts for 50.93%. The remaining variations are explained by the residuals, which is 51.34% in the J35 asymmetry model and 49.07% in the Jl asymmetry model.
The CC morphometric maps of the sub-groups and pooled samples are shown in Figure 5. Across all the CC morphometric maps, the SMA asymmetry values and the J35 or Jfifty asymmetry value are positively correlated amongst the entire humeral diaphysis, except for some areas of Henan. When specific to the morphometric maps of CC35, high CC35 values are detected primarily among the distal half of the diaphysis, especially around the mid-distal to distal department, while lower CC35 values are more than inclined to distribute anteroposteriorly over the proximal one-half of the diaphysis. Henan differs from the other sub-groups in that its SMA asymmetry values are negatively correlated with the J35 asymmetry value in the region between the mid-proximal and middle diaphysis, anteroposteriorly. For the morphometric maps of CCl, loftier CCl values are found betwixt the proximal and middle diaphysis, anteroposteriorly, which gradually shift in the anterolateral posteromedial attribute, from the middle to distal diaphysis. Comparatively, low CC50 values tend to follow the approximately anterolateral posteromedial aspect between the mid-distal and distal diaphysis. In comparing to other sub-groups, Henan exhibits a distinct distribution blueprint of CCl values at the distal humeral section, mediolaterally, with the SMA asymmetry values existence negatively correlated with the Jl disproportion value.
4. Discussion
The objective of this study was to reveal the humeral asymmetry patterns of East Asian modernistic humans with various backgrounds, by evaluating the biomechanical performance across complete humeral diaphysis rather than private cross-sections only, as well as to identify the reliability of torsional rigidity at the 35% and 50% cross-sections (J35 and Jfifty) in bilateral asymmetry analysis.
By quantifying the overall angle rigidity asymmetry of humeral proximodistal diaphysis using morphometric mapping of SMA asymmetry values, the variation range and pattern of humeral asymmetry in East Asian modern humans represented by our samples were investigated. In all the sub-groups, male humeri are more than asymmetrical than female humeri. The Henan population has lower humeral asymmetry overall compared to the Hubei and Xinjiang populations. Although three populations show unique distributions of bending rigidity asymmetry, the inter-grouping differences are not meaning in MANOVA. This suggests that, at least for the samples used in this study, the behavioral differences among unlike populations and between dissimilar sexes are not significant enough to generate discernable differences in bilateral asymmetry. The relatively small sample size of the present report might be a gene in this effect. Future studies with larger sample sizes and populations from more varied backgrounds may reveal significant differences.
Overall, the hateful morphometric maps of most the sub-groups and pooled samples show the following common distribution blueprint: the disproportion of the proximal department is reinforced anteroposteriorly, connecting it to some other relatively asymmetrical area betwixt the mid-proximal and heart diaphysis, mediolaterally, and finally extending to the distal end in the anterolateral posteromedial aspect. Previous inquiry found that humeral asymmetry was nearly prominent at the midshaft and decreased towards both the proximal and distal diaphyseal ends, and this blueprint can exist attributed to the general mechanical model that angle loads should be the greatest at mid-diaphyseal regions [55]. However, as revealed in the present study, the proximal to middle diaphysis tends to have a higher asymmetrical level than the distal one-half, and the differences tend to be more prominent among different anatomical directions than betwixt unlike sections along the humeral diaphysis. This asymmetry pattern emphasizes the necessity of examining multiple anatomical directions when analyzing bilateral asymmetry, and suggests that the mechanism regulating the response of the long bone to external stimuli might be more complicated than previously understood.
As some highly asymmetrical regions correspond with the positions of major muscle attachments, such as deltoid tuberosity and the crest of the greater tubercle [56], the distribution of areas with reinforced disproportion might reflect adaptions to muscle loadings, which were proved to be an important determinant of upper-limb strength [57,58,59]. In our report, factors such as genetic regulation and health condition can be excluded from the elements influencing the bilateral disproportion because the analysis was based on paired humeri from the aforementioned individual. Still, more than experimental evidences are needed to verify this hypothesis in future studies.
Co-ordinate to the results of the CV morphometric maps, the variability in bilateral asymmetry is not consistent across the humeral diaphysis. Highly variable regions are restricted to the distal half of humeral diaphysis in the anteromedial posterolateral aspect, corresponding to the medial/lateral edge and medial/lateral supracondylar. Since this feature is shared by all the sub-groups also as the pooled data, it may correspond a generality of East Asian modern humans. It is noteworthy that highly variable regions on the humeral diaphysis tend to overlap with areas presenting a low asymmetrical level, which may be a point of relative insensitivity to lateralized mechanical stimuli (see previous paragraph). Previous studies institute that humeral distal articular properties, such as articular expanse, did non but respond to mechanical loadings, just were also ontogenetically constrained and genetically canalized [60]. As the structure of the medial/lateral border and medial/lateral supracondylar are closely related to the distal articular morphology, co-ordinate to their anatomical adjacency [56], one possible interpretation for the loftier variability of disproportion is that these regions might present fluctuating asymmetry that is attributable to genetic, nutrient, and health factors instead of the mechanical surround solitary [lx,61,62].
This study supports the previous perspective that torsional rigidity at a specific cross-section (35% or 50% of the humeral biomechanical length) tin can be used to betoken the overall biomechanical disproportion of humeral diaphysis, because the multivariate regression model built on all the specimens is constructive, and a positive correlation exists between the SMA asymmetry and J asymmetry at most diaphyseal locations. However, nosotros should besides notation that a single J asymmetry value cannot convey the complexity of the entire humerus' asymmetry. The correlation betwixt overall SMA asymmetry and J asymmetry is moderate, considering J35 and J50 asymmetry can just explain about half of the total variation in humeral bilateral asymmetry. In addition, the degree of correlation betwixt SMA asymmetry and J asymmetry varies across the humeral diaphysis, and is but strong in specific regions.
5. Conclusions
This written report evaluated humeral biomechanical asymmetry across complete humeral diaphysis based on loftier-resolution micro-CT, and by quantifiable visualization and statistical methods. Using specimens from three Chinese archaeological populations that varied in geographic location, chronological age, and subsistence pattern, the pattern of humeral asymmetry in East Asian modern humans was investigated. Distinct humeral asymmetry patterns are observed on the mean morphometric map, but are not statistically pregnant. Coordinating distributions of highly asymmetrical regions and CV are observed across nearly all the sexes and populations, indicating possible universality of the humeral asymmetry design in East Asian modern humans. These highly asymmetrical regions stand for with major muscle attachments. The diaphyseal regions that are highly varied in bilateral asymmetry tend to present a low asymmetrical level. Although J35 and J50 asymmetry are related to the overall humeral asymmetry, it can simply explain near half of the full variation. These findings suggest that the overall biomechanical asymmetry of humeral diaphysis is more complicated than previously assumed. This report complements previous findings on humeral asymmetry, and accrue information and knowledge for futurity works in this surface area.
Writer Contributions
Conceptualization, Y.Z.; methodology, Y.Z.; software, Y.Z.; validation, Y.Z., S.Ten.; formal assay, Y.Z.; investigation, Y.Z.; resources, M.Z., H.L., J.H.; data curation, Y.Z., Southward.X.; writing—original draft preparation, Y.Z.; writing—review and editing, P.West., S.X.; visualization, Y.Z.; supervision, S.X.; project administration, S.X.; funding conquering, H.L., S.10. All authors have read and agreed to the published version of the manuscript.
Funding
This inquiry was funded by the Strategic Priority Inquiry Program of the Chinese Academy of Sciences (XDB26000000), the National Natural Science Foundation of Communist china (41872030), and the National Social Science Fund of Communist china (19BKG039, 19VJX066).
Institutional Review Board Argument
Not applicative.
Informed Consent Statement
Not applicable.
Information Availability Statement
The data supporting this report are available from the corresponding author on reasonable request.
Acknowledgments
The authors thank Yemao Hou, Pengfei Yin, and Jiawei Ma for their help in scanning and image processing. The authors too thank Xiujie Wu and Mackie O'Hara Ali for optimizing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no function in the pattern of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A
Figure A1. Morphometric maps exhibiting SMA asymmetry values of all specimens.
Effigy A1. Morphometric maps exhibiting SMA asymmetry values of all specimens.
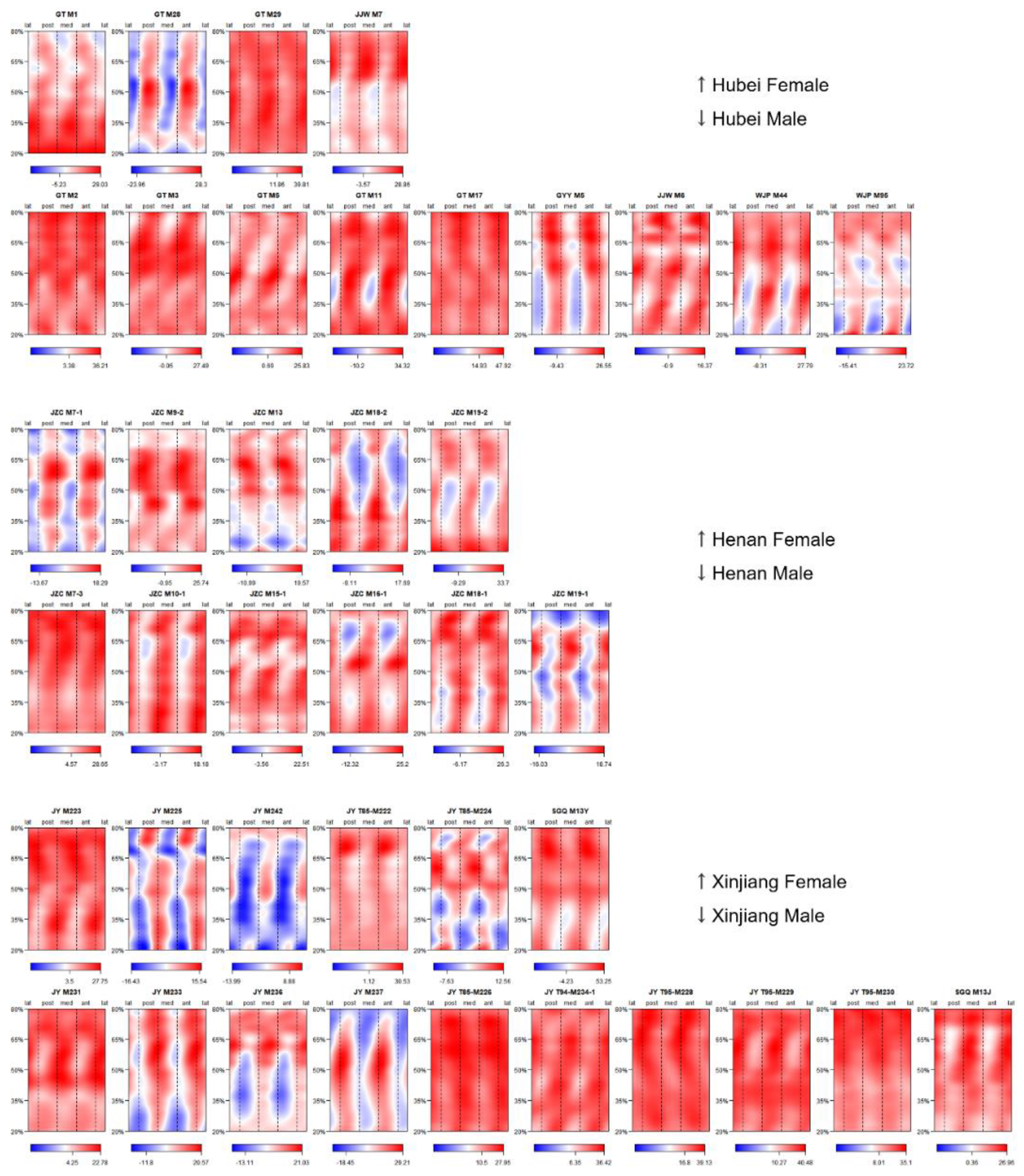
References
- DeLude, J.A.; Bicknell, R.T.; MacKenzie, Thou.A.; Ferreira, L.M.; Dunning, C.E.; Rex, One thousand.J.W.; Johnson, J.A.; Drosdowech, D.S. An anthropometric study of the bilateral anatomy of the humerus. J. Shoulder Elbow Surg. 2007, sixteen, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Ubelaker, D.H.; Zarenko, K.M. Tin handedness exist adamant from skeletal remains? A chronological review of the literature. J. Forensic Sci. 2012, 57, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Ruff, C.B. Biomechanical analyses of archaeological human skeletons. In Biological Anthropology of the Homo Skeleton, 3rd ed.; Katzenberg, M.A., Grauer, A.L., Eds.; Wiley-Blackwell: Hoboken, NJ, Usa, 2018; pp. 189–224. [Google Scholar]
- Faurie, C.; Raymond, M. Handedness, homicide and negative frequency-dependent choice. Proc. R. Soc. B 2005, 272, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.N. Is 'mitt preference' coded in the hominin skeleton? An in-vivo written report of bilateral morphological variation. J. Hum. Evol. 2011, 61, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Ozener, B. Extreme behavioral lateralization and the remodeling of the distal humerus. Am. J. Hum. Biol. 2012, 24, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Haapasalo, H.; Kannus, P.; Sievänen, H.; Pasanen, 1000.; Uusi-Rasi, Thousand.; Heinonen, A.; Oja, P.; Vuori, I. Event of long-term unilateral activity on bone mineral density of female person junior tennis players. J. Os Miner. Res. 1998, 13, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Bass, South.L.; Saxon, L.; Daly, R.M.; Turner, C.H.; Robling, A.K.; Seeman, East.; Stuckey, S. The consequence of mechanical loading on the size and shape of bone in pre-, peri-, and postpubertal girls: A study in tennis players. J. Bone Miner. Res. 2002, 17, 2274–2280. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.N.; Stock, J.T. Habitual throwing and swimming represent with upper limb diaphyseal force and shape in modernistic human being athletes. Am. J. Phys. Anthropol. 2009, 140, 160–172. [Google Scholar] [CrossRef]
- Balzeau, A.; Brawl-Albessard, L.; Kubicka, A.Grand. Variation and correlations in departures from symmetry of brain torque, humeral morphology and handedness in an archaeological sample of Homo sapiens. Symmetry 2020, 12, 432. [Google Scholar] [CrossRef]
- Shackelford, Fifty.L. Regional variation in the postcranial robusticity of belatedly upper paleolithic humans. Am. J. Phys. Anthropol. 2007, 133, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Sparacello, V.S.; d'Ercole, V.; Coppa, A. A bioarchaeological approach to the reconstruction of changes in military organization amidst Iron Historic period Samnites (Vestini) From Abruzzo, Central Italy. Am. J. Phys. Anthropol. 2015, 156, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Sládek, V.; Ruff, C.B.; Berner, M.; Holt, B.; Niskanen, M.; Schuplerová, E.; Hora, M. The impact of subsistence changes on humeral bilateral asymmetry in Terminal Pleistocene and Holocene Europe. J. Hum. Evol. 2016, 92, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Sparacello, Five.South.; Villotte, S.; Shackelford, Fifty.L.; Trinkaus, E. Patterns of humeral asymmetry among Late Pleistocene humans. Comptes Rendus Palevol 2017, 16, 680–689. [Google Scholar] [CrossRef]
- Wei, P.; Lu, H.; Carlson, K.J.; Zhang, H.; Hui, J.; Zhu, M.; He, K.; Jashashvili, T.; Zhang, X.; Yuan, H.; et al. The upper limb skeleton and behavioral lateralization of modern humans from Zhaoguo Cave, southwestern China. Am. J. Phys. Anthropol. 2020, 173, 671–696. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Zhao, Y.; Walker, C.S.; He, J.; Lu, X.; Hui, J.; Shui, W.; Jin, L.; Liu, W. Internal structural properties of the humeral diaphyses in an early modern man from Tianyuan Cave, Mainland china. Quat. Int. 2021, 591, 107–118. [Google Scholar] [CrossRef]
- Hsieh, Y.-F.; Robling, A.M.; Ambrosius, W.T.; Burr, D.B.; Turner, C.H. Mechanical loading of diaphyseal os in vivo: The strain threshold for an osteogenic response varies with location. J. Bone Miner. Res. 2001, 16, 2291–2297. [Google Scholar] [CrossRef] [PubMed]
- Burr, D.B.; Robling, A.K.; Turner, C.H. Effects of biomechanical stress on bones in animals. Bone 2002, 30, 781–786. [Google Scholar] [CrossRef]
- Pearson, O.Yard.; Lieberman, D.E. The aging of Wolff's "constabulary": Ontogeny and responses to mechanical loading in cortical bone. Am. J. Phys. Anthropol. 2004, 125, 63–99. [Google Scholar] [CrossRef] [PubMed]
- Ruff, C.; Holt, B.; Trinkaus, E. Who's afraid of the large bad wolff? "Wolff is law" and bone functional accommodation. Am. J. Phys. Anthropol. 2006, 129, 484–498. [Google Scholar] [CrossRef] [PubMed]
- Gosman, J.H.; Hubbell, Z.R.; Shaw, C.North.; Ryan, T.One thousand. Development of cortical bone geometry in the human femoral and tibial diaphysis. Anat. Rec. 2013, 296, 774–787. [Google Scholar] [CrossRef]
- Sládek, V.; Berner, Grand.; Holt, B.; Niskanen, M.; Ruff, C.B. Past man manipulative beliefs in the European Holocene as assessed through upper limb asymmetry. In Skeletal Variation and Accommodation in Europeans; Wiley-Blackwell: Hoboken, NJ, United states of america, 2018; pp. 163–208. [Google Scholar]
- Perchalski, B.; Placke, A.; Sukhdeo, S.M.; Shaw, C.N.; Gosman, J.H.; Raichlen, D.A.; Ryan, T.Thousand. Asymmetry in the cortical and trabecular bone of the human humerus during development. Anat. Rec. 2018, 301, 1012–1025. [Google Scholar] [CrossRef] [PubMed]
- Hong, East.; Kwak, D.-S.; Kim, I.-B. Morphometric evaluation of detailed asymmetry for the proximal humerus in Korean population. Symmetry 2021, 13, 862. [Google Scholar] [CrossRef]
- Ruff, C.B.; Burgess, M.L.; Bromage, T.G.; Mudakikwa, A.; McFarlin, S.C. Ontogenetic changes in limb bone structural proportions in mount gorillas (Gorilla beringei beringei). J. Hum. Evol. 2013, 65, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Ruff, C.B. Long bone articular and diaphyseal structure in erstwhile world monkeys and apes. I: Locomotor effects. Am. J. Phys. Anthropol. 2002, 119, 305–342. [Google Scholar] [CrossRef] [PubMed]
- Weiss, E. Humeral cantankerous-sectional morphology from 18th century Quebec prisoners of war: Limits to activity reconstruction. Am. J. Phys. Anthropol. 2005, 126, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Wescott, D.J.; Cunningham, D.L. Temporal changes in Arikara humeral and femoral cross-sectional geometry associated with horticultural intensification. J. Archaeol. Sci. 2006, 33, 1022–1036. [Google Scholar] [CrossRef]
- Sládek, Five.; Berner, Grand.; Sosna, D.; Sailer, R. Homo manipulative behavior in the Central European Late Eneolithic and Early Bronze Age: Humeral bilateral asymmetry. Am. J. Phys. Anthropol. 2007, 133, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.A.; Knüsel, C.J. Activity-related skeletal alter in medieval humeri: Cross-exclusive and architectural alterations. Am. J. Phys. Anthropol. 2005, 128, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.C.; Pearson, O.Thousand.; Durband, A.C.; Walshe, One thousand.; Carlson, K.J.; Grine, F.Eastward. An examination of the cross-sectional geometrical backdrop of the long bone diaphyses of Holocene foragers from Roonka, South Australia. Am. J. Phys. Anthropol. 2020, 172, 682–697. [Google Scholar] [CrossRef] [PubMed]
- Cowgill, L.W.; Mednikova, M.B.; Buzhilova, A.P.; Trinkaus, Eastward. The Sunghir 3 Upper Paleolithic juvenile: Pathology versus persistence in the Paleolithic. Int. J. Osteoarchaeol. 2015, 25, 176–187. [Google Scholar] [CrossRef]
- Shaw, C.Northward.; Stock, J.T. Extreme mobility in the Belatedly Pleistocene? Comparing limb biomechanics among fossil Homo, varsity athletes and Holocene foragers. J. Hum. Evol. 2013, 64, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Trinkaus, E. Epipaleolithic human being appendicular remains from Ein Gev I, Israel. Comptes Rendus Palevol 2018, 17, 616–627. [Google Scholar] [CrossRef]
- Kubicka, A.M.; Nowaczewska, West.; Balzeau, A.; Piontek, J. Bilateral asymmetry of the humerus in Neandertals, Australian aborigines and medieval humans. Am. J. Phys. Anthropol. 2018, 167, 46–60. [Google Scholar] [CrossRef]
- Ogilvie, Yard.D.; Hilton, C.E. Cantankerous-sectional geometry in the humeri of foragers and farmers from the prehispanic American Southwest: Exploring patterns in the sexual sectionalisation of labor. Am. J. Phys. Anthropol. 2011, 144, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Sparacello, V.Due south.; Pearson, O.M.; Coppa, A.; Marchi, D. Changes in skeletal robusticity in an iron historic period agropastoral group: The samnites from the Alfedena necropolis (Abruzzo, Central Italy). Am. J. Phys. Anthropol. 2011, 144, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Warden, Due south.J.; Mantila Roosa, S.M.; Kersh, M.Eastward.; Hurd, A.L.; Fleisig, 1000.Southward.; Pandy, M.G.; Fuchs, R.K. Physical activeness when immature provides lifelong benefits to cortical bone size and forcefulness in men. Proc. Natl. Acad. Sci. Us 2014, 111, 5337–5342. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.A.B.; Humphrey, L.T. A Virtual geometric morphometric approach to the quantification of long bone bilateral disproportion and cross-sectional shape. Am. J. Phys. Anthropol. 2015, 158, 541–556. [Google Scholar] [CrossRef]
- Bondioli, L.; Bayle, P.; Dean, C.; Mazurier, A.; Puymerail, L.; Ruff, C.; Stock, J.T.; Volpato, V.; Zanolli, C.; Macchiarelli, R. Technical notation: Morphometric maps of long bone shafts and dental roots for imaging topographic thickness variation. Am. J. Phys. Anthropol. 2010, 142, 328–334. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhou, Yard.; Wei, P.; Xing, S. second visualization and quantitative analysis of the humeral diaphysis cortical thickness. Acta Anthropol. Sin. 2020, 39, 632–647. [Google Scholar] [CrossRef]
- Puymerail, L.; Ruff, C.B.; Bondioli, L.; Widianto, H.; Trinkaus, Eastward.; Macchiarelli, R. Structural analysis of the Kresna 11 Homo erectus femoral shaft (Sangiran, Java). J. Hum. Evol. 2012, 63, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Puymerail, L.; Volpato, V.; Debénath, A.; Mazurier, A.; Tournepiche, J.-F.; Macchiarelli, R. A Neanderthal partial femoral diaphysis from the "grotte de la Bout", La Chaise-de-Vouthon (Charente, France): Outer morphology and endostructural arrangement. Comptes Rendus Palevol 2012, xi, 581–593. [Google Scholar] [CrossRef]
- Morimoto, North.; Ponce de León, G.South.; Zollikofer, C.P.E. Exploring femoral diaphyseal shape variation in wild and captive chimpanzees by means of morphometric mapping: A test of Wolff'south law. Anat. Rec. 2011, 294, 589–609. [Google Scholar] [CrossRef] [PubMed]
- Jashashvili, T.; Dowdeswell, 1000.R.; Lebrun, R.; Carlson, 1000.J. Cortical structure of hallucal metatarsals and locomotor adaptations in hominoids. PLoS ONE 2015, ten, e0117905. [Google Scholar] [CrossRef]
- Cameron, 1000.East.; Pfeiffer, S. Long bone cross-sectional geometric properties of Later Stone Historic period foragers and herder-foragers. South. Afr. J. Sci. 2014, 110, 01–11. [Google Scholar] [CrossRef]
- Tian, J.; Tang, L.; Shi, D.; Luo, Y.; Zhao, Z. Research on the charred institute remains unearthed from the Jijiawan site in Fang County, Hubei Province. Cult. Relics South. China 2019, v, 180–188. (In Chinese) [Google Scholar] [CrossRef]
- Xing, S.; Carlson, Yard.J.; Wei, P.; He, J.; Liu, Due west. Morphology and structure of Human being erectus humeri from Zhoukoudian, Locality ane. PeerJ 2018, 6, e4279. [Google Scholar] [CrossRef] [PubMed]
- Wang, 50.; Xiao, M.; Liu, Z.; Lv, Due east.; Wu, Y. Cursory report on digging of Jiayi cemeteries in Turpan, Xinjiang. Turfanological Res. 2014, 1, 1–nineteen. [Google Scholar] [CrossRef]
- Xing, K.; Zhang, Y.; Li, Z. 新疆鄯善三个桥墓葬发掘简报. Cult. Relics 2002, 6, 46–56. (In Chinese). Available online: http://www.cqvip.com/qk/97337x/2002006/11400528.html (accessed on 27 Baronial 2021).
- Xiao, One thousand. The Review and Written report of Jia Yi Cemetery in Turfan, Xinjiang. Master's Thesis, Northwest Academy, Xi'an, People's republic of china, 2018. [Google Scholar]
- Profico, A.; Bondioli, Fifty.; Raia, P.; O'Higgins, P.; Marchi, D. morphomap: An R bundle for long bone landmarking, cortical thickness, and cantankerous-exclusive geometry mapping. Am. J. Phys. Anthropol. 2021, 174, 129–139. [Google Scholar] [CrossRef]
- Collyer, 1000.L.; Adams, D.C. RRPP: An r package for fitting linear models to high-dimensional data using balance randomization. Methods Ecol. Evol. 2018, 9, 1772–1779. [Google Scholar] [CrossRef]
- Adams, D.C.; Otárola-Castillo, E. geomorph: An r bundle for the drove and assay of geometric morphometric shape data. Methods Ecol. Evol. 2013, 4, 393–399. [Google Scholar] [CrossRef]
- Zelazny, M.One thousand.; Sylvester, A.D.; Ruff, C.B. Bilateral asymmetry and developmental plasticity of the humerus in modern humans. Am. J. Phys. Anthropol. 2021, 174, 418–433. [Google Scholar] [CrossRef] [PubMed]
- White, T.D.; Black, M.T.; Folkens, P.A. Arm: Humerus, Radius, and Ulna. In Human being Osteology, 3rd ed.; Academic Press: San Diego, CA, USA, 2012; pp. 175–198. [Google Scholar]
- Ruff, C. Growth in bone strength, body size, and muscle size in a juvenile longitudinal sample. Os 2003, 33, 317–329. [Google Scholar] [CrossRef]
- Schoenau, Eastward.; Neu, C.M.; Mokov, E.; Wassmer, G.; Manz, F. Influence of puberty on musculus area and cortical bone expanse of the forearm in boys and girls. J. Clin. Endocrinol. Metab. 2000, 85, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Niinimäki, Southward. The relationship between musculoskeletal stress markers and biomechanical properties of the humeral diaphysis. Am. J. Phys. Anthropol. 2012, 147, 618–628. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, D.East.; Devlin, M.J.; Pearson, O.Yard. Articular area responses to mechanical loading: Furnishings of exercise, age, and skeletal location. Am. J. Phys. Anthropol. 2001, 116, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Plochocki, J.H. Bilateral variation in limb articular surface dimensions. Am. J. Hum. Biol. 2004, 16, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Hallgrímsson, B.; Willmore, K.; Hall, B.Thousand. Canalization, developmental stability, and morphological integration in primate limbs. Am. J. Phys. Anthropol. 2002, 119, 131–158. [Google Scholar] [CrossRef]
Figure 1. The positional and directional correspondence between humeral external structure, diaphyseal cross-sections, and morphometric map exhibiting bending rigidity disproportion. Abbreviations for anatomical terms are as follows: prox: proximal; mid: middle; dist: distal; lat: lateral; mail service: posterior; med: medial; ant: inductive.
Figure i. The positional and directional correspondence between humeral external structure, diaphyseal cross-sections, and morphometric map exhibiting bending rigidity asymmetry. Abbreviations for anatomical terms are as follows: prox: proximal; mid: middle; dist: distal; lat: lateral; post: posterior; med: medial; pismire: anterior.
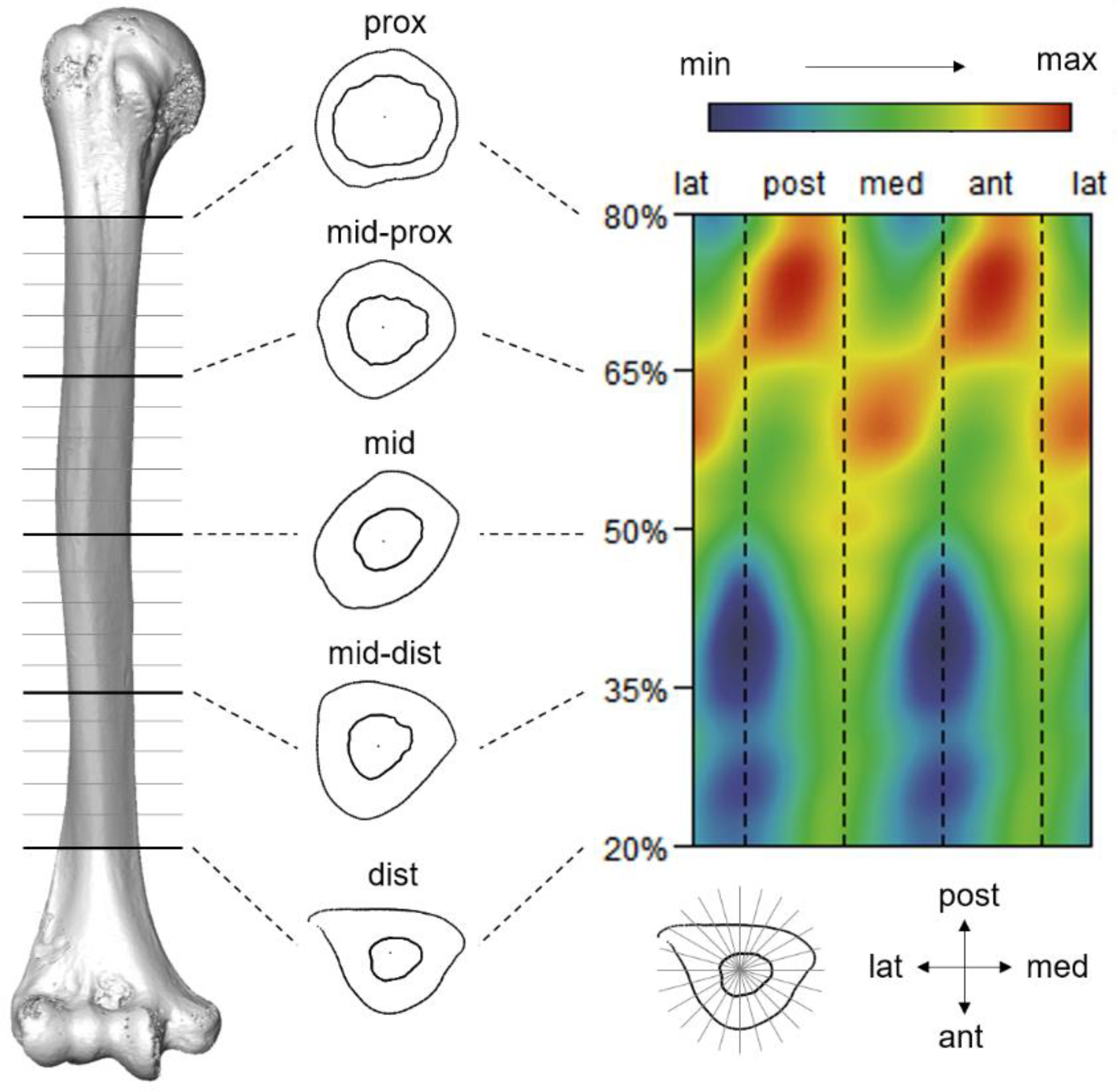
Figure 2. Mean morphometric maps exhibiting SMA disproportion values for sub-groups and pooled samples (P). Sub-groups are defined by population, sexual practice, and the pairwise combination of these 2 factors. Populations include Hubei (HB), Henan (HN), and Xinjiang (XJ); sexes include female (F) and male (M). All mean morphometric maps are under the same chromatic scale.
Figure 2. Mean morphometric maps exhibiting SMA disproportion values for sub-groups and pooled samples (P). Sub-groups are divers past population, sex, and the pairwise combination of these ii factors. Populations include Hubei (HB), Henan (HN), and Xinjiang (XJ); sexes include female (F) and male (Yard). All mean morphometric maps are nether the same chromatic scale.
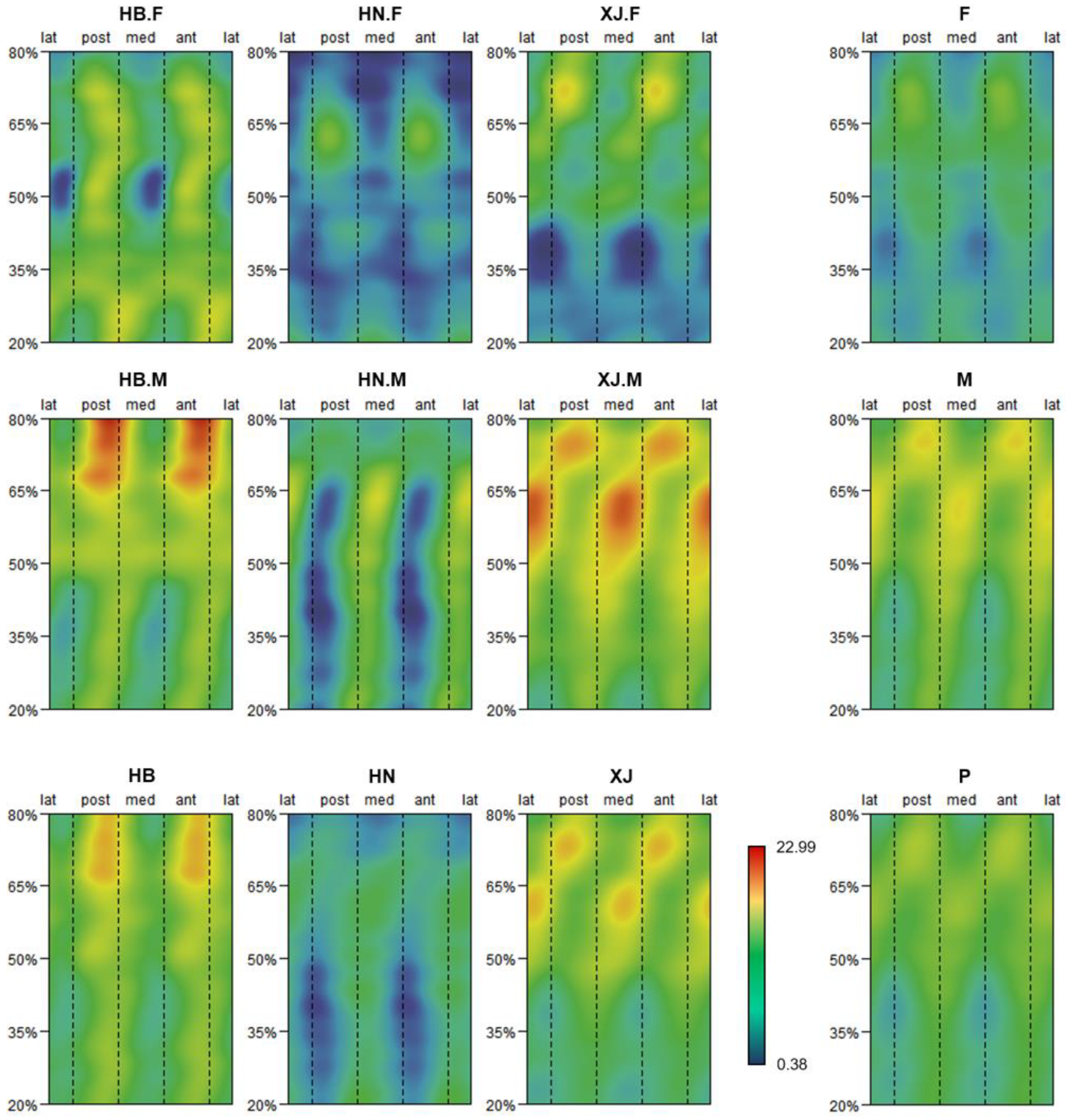
Figure 3. Morphometric maps exhibiting the coefficient of variation (CV) for SMA asymmetry values in sub-groups and pooled samples (P). Sub-groups are defined by population and sexual activity. Populations include Hubei (HB), Henan (HN), and Xinjiang (XJ); sexes include female (F) and male (M). All CV morphometric maps are under the same chromatic scale.
Figure 3. Morphometric maps exhibiting the coefficient of variation (CV) for SMA disproportion values in sub-groups and pooled samples (P). Sub-groups are defined by population and sex. Populations include Hubei (HB), Henan (HN), and Xinjiang (XJ); sexes include female person (F) and male (Chiliad). All CV morphometric maps are under the same chromatic scale.
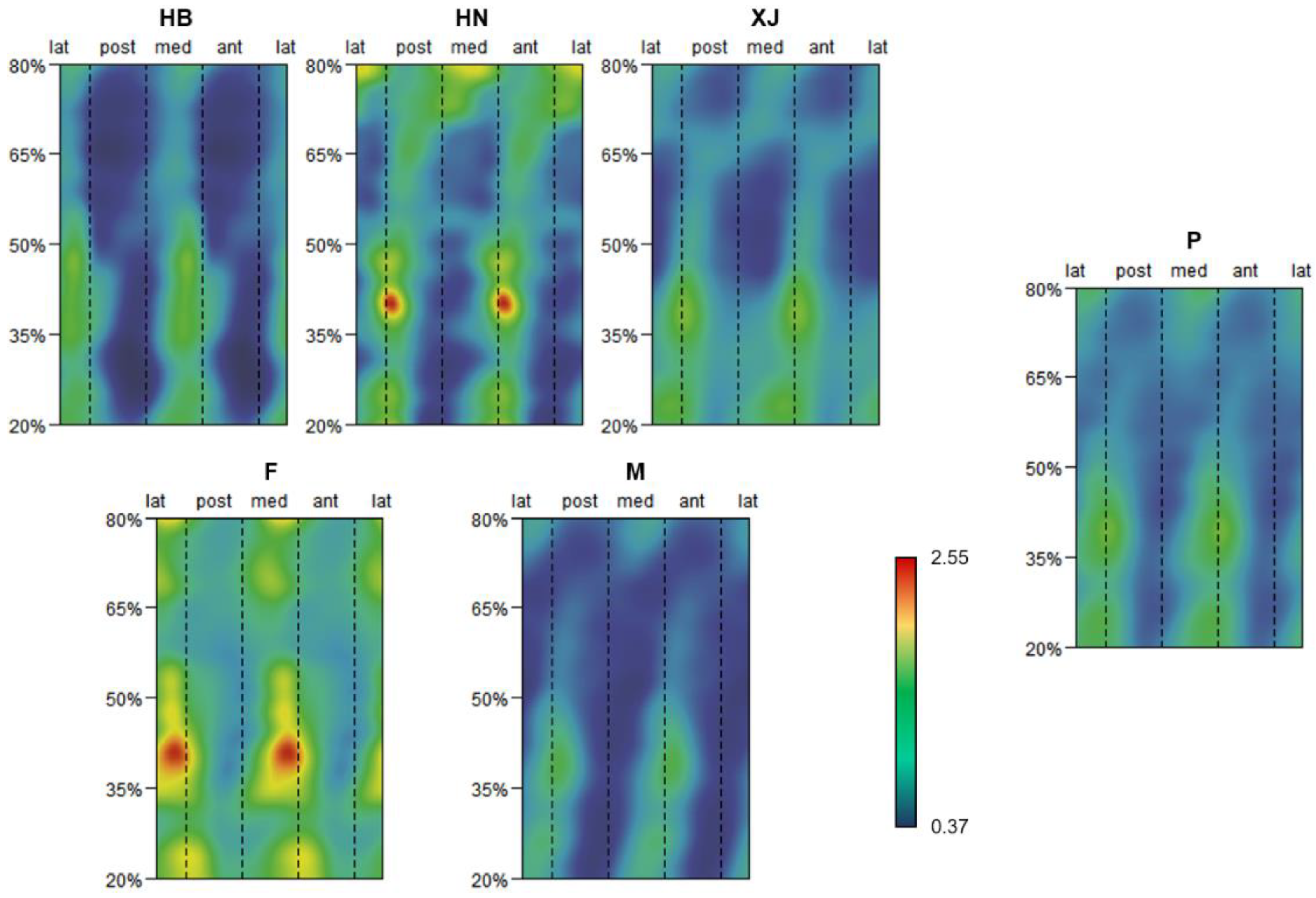
Figure four. Multivariate regression of all SMA asymmetry values on J asymmetry value at the 35% or 50% cross-section using pooled samples.
Figure iv. Multivariate regression of all SMA asymmetry values on J asymmetry value at the 35% or l% cross-section using pooled samples.
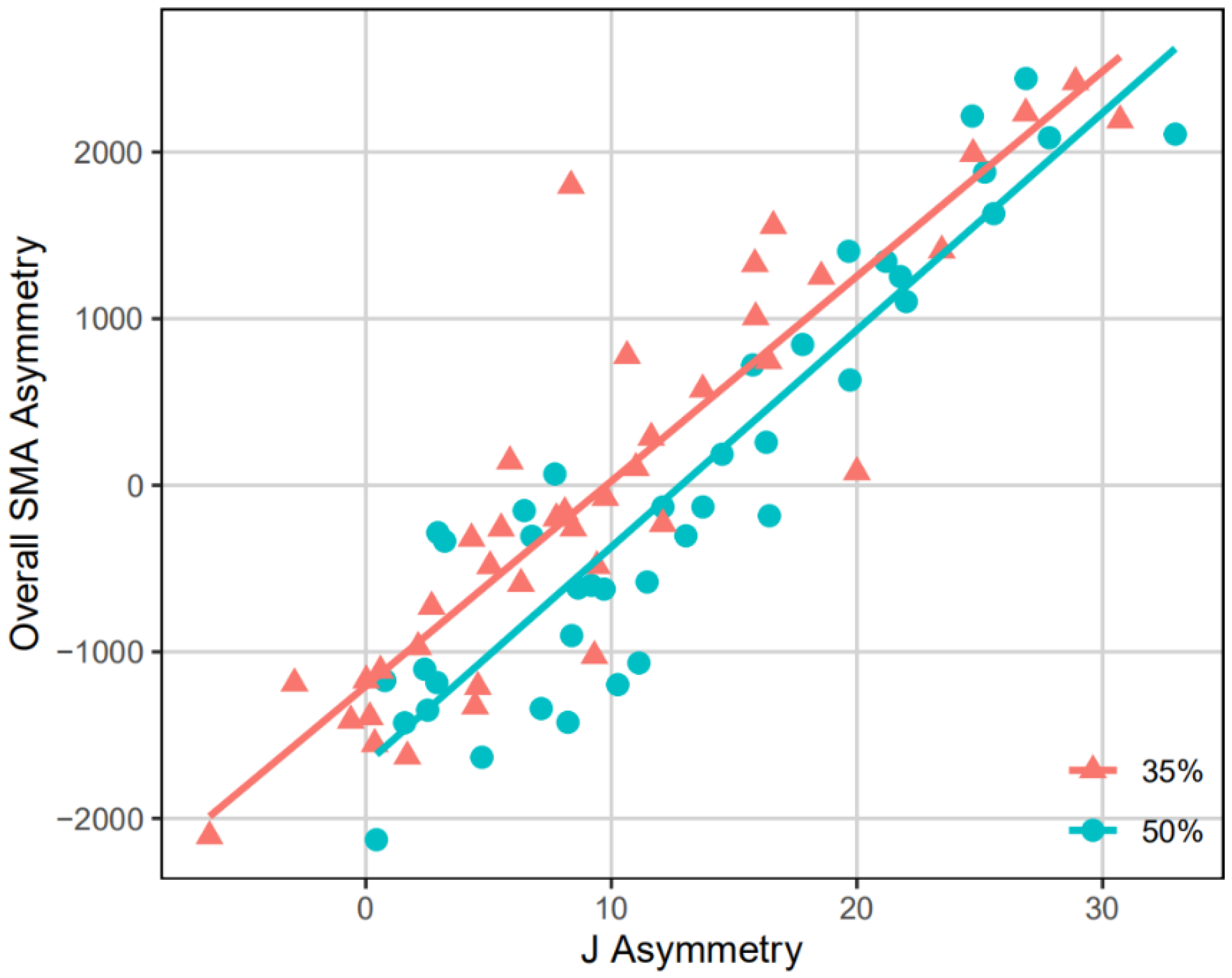
Figure 5. Morphometric maps exhibiting the correlation coefficient (CC) between SMA disproportion values and J disproportion value at the 35% or 50% cross-section in sub-groups and pooled samples (P). Sub-groups are defined by population and sex. Populations include Hubei (HB), Henan (HN), and Xinjiang (XJ); sexes include female (F) and male (1000). All CC morphometric maps are under the same chromatic scale.
Figure 5. Morphometric maps exhibiting the correlation coefficient (CC) between SMA asymmetry values and J disproportion value at the 35% or 50% cross-department in sub-groups and pooled samples (P). Sub-groups are defined by population and sex. Populations include Hubei (HB), Henan (HN), and Xinjiang (XJ); sexes include female person (F) and male (M). All CC morphometric maps are under the same chromatic scale.
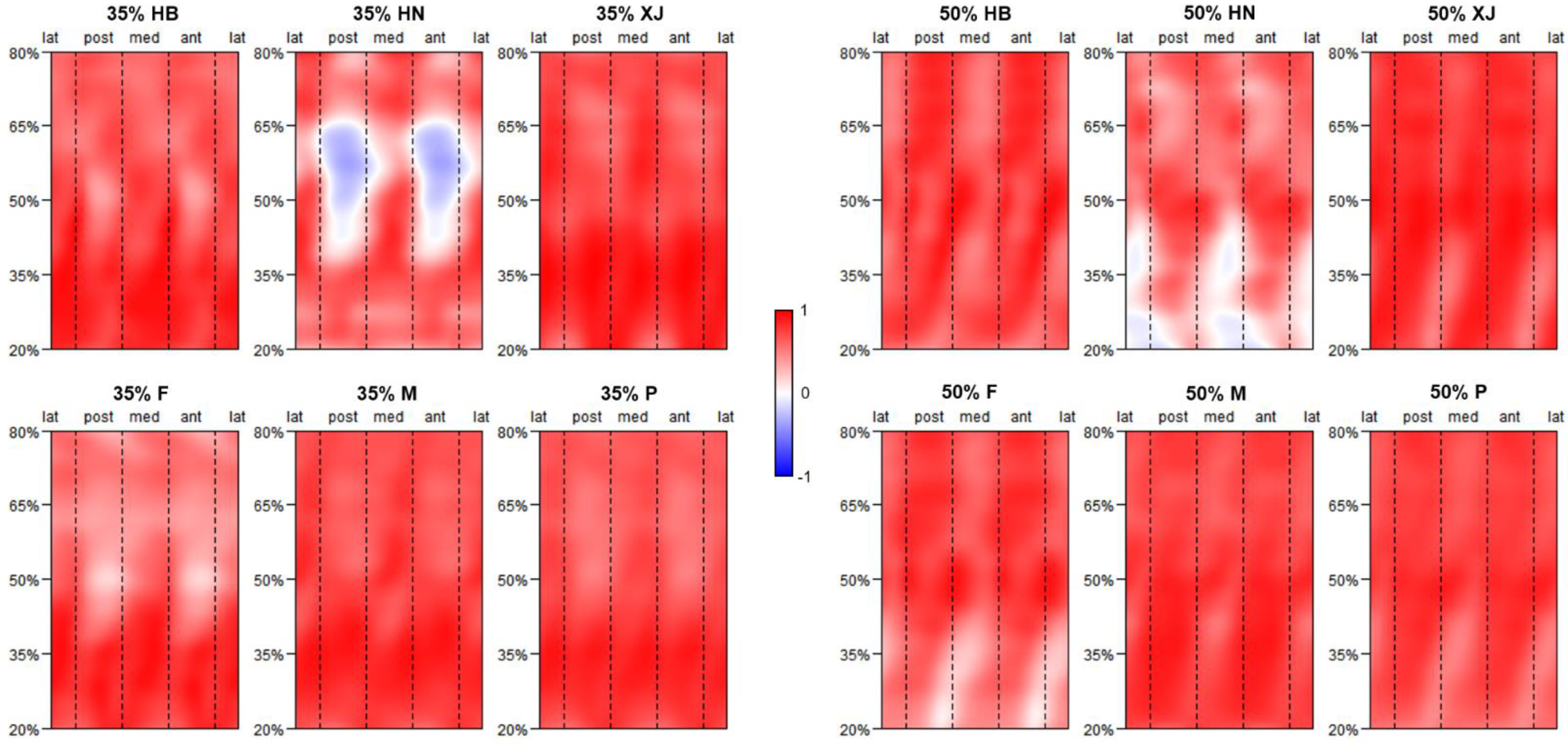
Table ane. MANOVA results interpreting the differences betwixt sexes and among populations.
Table 1. MANOVA results interpreting the differences between sexes and amidst populations.
| Df | SS | MS | Rsq | F | Z | P (>F) | |
|---|---|---|---|---|---|---|---|
| Sexual practice | 1 | 5,309,415 | 5,309,415 | 0.05494 | 2.1778 | 1.31140 | 0.110 |
| Population | 2 | 5,792,218 | 2,896,109 | 0.05993 | one.1879 | 0.58014 | 0.296 |
| Sexual activity:Population | 2 | ii,652,077 | one,326,039 | 0.02744 | 0.5439 | −0.78875 | 0.783 |
| Residuals | 34 | 82,891,403 | 2,437,982 | 0.85769 | |||
| Full | 39 | 96,645,113 |
Table 2. Multivariate regression of all SMA asymmetry values on J asymmetry value.
Table 2. Multivariate regression of all SMA disproportion values on J asymmetry value.
| Df | SS | MS | Rsq | F | Z | P (>F) | |
|---|---|---|---|---|---|---|---|
| J35 asymmetry | 1 | 47,026,558 | 47,026,558 | 0.48659 | 36.015 | iii.5479 | 0.001 ** |
| Residuals | 38 | 49,618,555 | ane,305,751 | 0.51341 | |||
| Total | 39 | 96,645,113 | |||||
| J50 disproportion | 1 | 49,218,068 | 49,218,068 | 0.50927 | 39.435 | 3.7475 | 0.001 ** |
| Residuals | 38 | 47,427,046 | one,248,080 | 0.49073 | |||
| Full | 39 | 96,645,113 |
| Publisher's Notation: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and weather condition of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/four.0/).
Source: https://www.mdpi.com/2073-8994/13/10/1843/htm
0 Response to "Can Handedness Be Determined From Skeletal Remains? A Chronological Review of the Literature"
Post a Comment